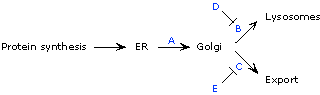Remember that a mutation in the last gene in the pathway can only be rescued by the final product; a mutation in the next-to-last gene can be rescued by the last two compounds in the pathway, etc. Thus, the pathway is:
![]()
Answer key, practice problems 2000--week 8
| 1. | (a) | Because the two mutant strains showed complementation (the F1 were able to hear), the mutations must have been in separate genes; the simplest explanation is that there are two genes involved. This conclusion is supported by the F2 ratio, which can be derived from a dihybrid ratio. | ||||||||
| (b) |
The 9:7 F2 ratio indicates that we are dealing with a dihybrid ratio (the fractions go in sixteenths). The 9:7 ratio can be derived from the standard 9:3:3:1 ratio if we postulate that the 9/16 of progeny that have at least one dominant allele for each gene show the dominant phenotype (hearing) while the remaining 7/16 are homozygous recessive for at least one of the genes show the recessive phenotype (deafness). We would conclude then that the dominant phenotype requires that at least one dominant allele be present at each locus. If we call the two genes D and E, the parents were ddEE and DDee; the F1 progeny are DdEe (and can therefore hear); the F2 progeny are:
If the F1 rats were crossed to homozygous recessive rats (i.e., DdEe x ddee) the progeny would be DdEe, Ddee, ddEe, and ddee in equal proportions -- i.e., 1/4 of the progeny will be able to hear, and 3/4 will be deaf. |
|||||||||
| (c) | As with any dihybrid cross, one quarter of the progeny will be true breeding. |
| 2. | A selection for lacZ+ revertants: grow cells on non-lactose medium (to get some cells to work with), then plate them on agar plates containing lactose as the sole carbon source. Only lacZ+ revertants will be able to grow and form colonies. Therefore, any E. coli colonies that form on these plates must have a functional lacZ gene. |
| A screen: grow the cells as before, and plate them on non-lactose medium containing X-gal. All cells (lacZ- and lacZ+ revertants) will be able to grow, but only the lacZ+ revertants will be able to cleave X-gal to make blue colonies. |
| 3. |
|
| 4. | (a) | The genotype has to be A_H_ to show the A blood type; a genotype with either OO or hh -- or both, of course -- will give O blood type. Therefore, the progeny will be 9/16 A: 7/16 O blood type. (Draw a Punnett square if you are confused.) |
| (b) | 3/16 AB : 3/16 A : 3/16 B : 7/16 O | |
| (c) |
|
| 5. | (a) |
As with any independently assorting pair of genes,we can look at the the ratios for the two genes independently. With respect to presence or absence of color (the E gene), the progeny are 1/2 colored (black or brown) and half yellow. Therefore, with respect to the E gene, the parents were Ee and ee. With respect to black vs. brown (the B gene), the brown parent has to be bb and the other parent must be Bb (there must be at least one B allele to give black progeny; it cannot be BB, or there would be no brown progeny). [Another way to think about this is that of the non-yellow progeny, half are brown and half black; therefore, the parents must be bb and Bb giving 1:1 B_ and bb progeny.] Thus, the parents must be bbEe (brown) and Bbee (yellow). |
| (b) | Here, with respect to the E gene, one quarter of the progeny show the homozygous recessive phenotype--therefore both parents must be heterozygous (Ee) for the E gene. WIth respect to the B gene, again, black and brown progeny are in equal proportions, so the parents must be Bb and bb. Once again, the brown parent must be bbEe; the black parent must be BbEe. |
| 6. | (a) | D and E will rescue (because if either one is provided, E3 function will no longer be needed); C will accumulate (because there is no E3 to convert C to D). |
| (b) | E will rescue; D will accumulate | |
| (c) | D and E will rescue; B will accumulate. |
| 7. | (a) | Conversion of B to D cannot proceed, so D and F will each rescue this mutation. |
| (b) | Conversion of A to B cannot proceed, so B will rescue. |
| 8. | (a) | Red pigment cannot be made, so the flowers will be blue. |
| (b) | Red flowers. | |
| (c) | Purple flowers (because of complementation-- the F1 will be heterozygous for each gene). | |
| (d) | 9/16 purple: 3/16 red: 3/16 blue: 1/16 white. |
| 9. |
Remember that a mutation in the last gene in the pathway can only be rescued by the final product; a mutation in the next-to-last gene can be rescued by the last two compounds in the pathway, etc. Thus, the pathway is:
|
|
| 10. | (a) | The third cross alone shows evidence of two genes interacting, as it gives F2 ratios in sixteenths, as would be expected for a dihybrid cross. The ratio (9:3:4) is typical of recessive epistasis. | ||||||||||||
| (b) |
The first and the second cross show a 3:1 ratio in the F2 progeny, which looks like a monohybrid cross. However, we know that there are two genes contributing to flower color. So the obvious explanation is that in each of these crosses, one locus is homozygous for the same allele in both parents, so only one locus contributes to the observed F2 ratio. In the first cross, the white parent must be RRhh and the fuchsia parent must be homozygous normal RRHH. That would give RRHh F1 plants, and RRH_ and RRhh plants in 3:1 ratio in F2. By the same logic, the red parent in the second cross must be rrHH, while the fuchsia parent is RRHH again; the F2 are R_HH and rrHH in 3:1 ratio. In the third cross, the parents are RRhh and rrHH.
|
|||||||||||||
| (c) |
Since both genes are required for fuchsia color, and the white gene is epistatic to the red gene, the H gene must act earlier in the pathway, converting a colorless (white) precursor to red, while the R gene converts this red intermediate to fuchsia:
|
| 11. | The protein can clearly take two paths, either to the lysosome or getting exported from the cell. In the absence of gene A, neither pathway functions; instead, the protein accumulates in the ER. This result suggests that gene A functions early in the pathway, and is required to move the protein out of the ER. (Note: in this explanation, "gene A" can be taken to mean "product of gene A", etc., unless otherwise stated.) Gene B is needed to send the protein into lysosomes (because in the absence of gene B product, the protein is exported only); likewise, gene C is needed for export of the protein. So thus far, we surmise that gene A sends the protein out of the ER and that subsequently, gene B sends it into lysosomes while gene C exports it. This hypothesis is confirmed by the phenotype of A-B- and A-C- mutants, which show that A is epistatic to B and C. The B-C- double mutant phenotype shows that protein that gets past the ER but doesn't get into lysosomes or get exported ends up in the golgi--so the the function of A must be to transport the protein from the ER to the golgi; B and C transport the protein from the golgi to the lysosome or export the protein. |
|
What about genes D and E? In the absence of gene D, the lysosomal pathway seems to become overactive. Therefore, the function of gene D appears to be opposite that of gene B, suggesting that it is a repressor of gene B. Consistent with this hypothesis, B is epistatic to D (in the absence of B and D, we see the B- phenotype). Likewise, in the absence of gene E, the export pathway becomes overactive; and C is epistatic to E, suggesting that E is a repressor of C. In the absence of D and E, neither pathway is repressed, so the balance is restored (although that need not be predicted). The completed pathway, consistent with all the mutant phenotypes, is:
(An alternative interpretation, the D activates C and that E activates B, is also consistent with the data, provided that B and C can be transcribed in the absence of E and D --i.e., that B and C by themselves show some basal level of transcription that is further increased by E and D.) |