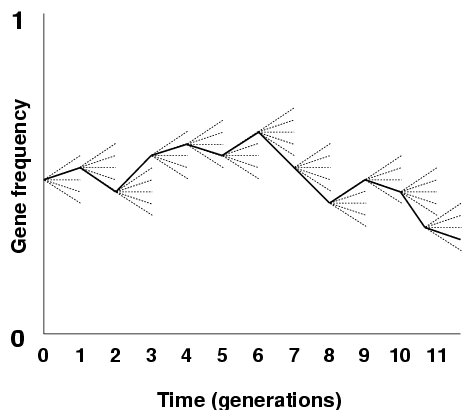
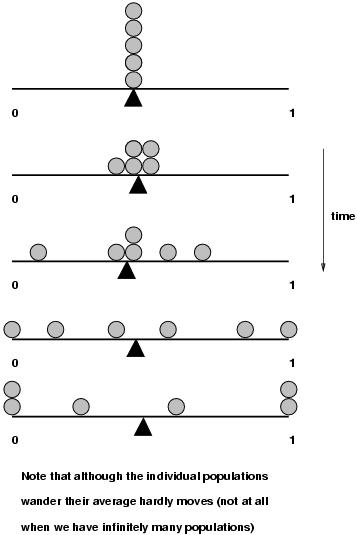
Mutation changes the genetic material in a cell: we will focus on mutations in the germ line. Mutations can arise spontanously (misrepair of DNA, or misreading of nucleotides) but this spontanous mutation rate can be increased through damage of the DNA by environmental factors, such as ultraviolet light, irradiation, and chemicals. Mutation rate is normally given in events per year or generation for a given a gene or a site on the DNA.

If we consider a locus with two possible alleles (A and a)) then we can consider a forward (u) and backward mutation (v). Forward mutation is the mutation from wildtype allele to the detrimental allele. Backward mutations undo the forward mutation. Because there are many ways to destroy the function but fewer ways to undo that harm, backward mutations are normally more rare than forward mutations.
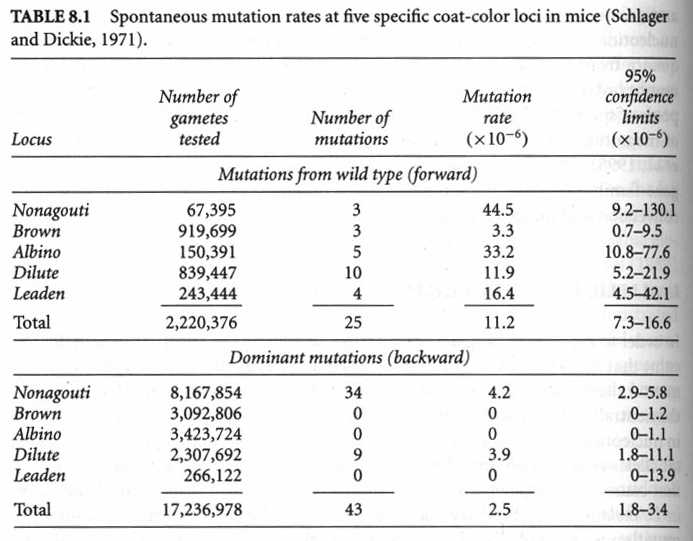
The frequency of A (p) in the next generation is
p1=p0(1-u) + (1-p0) v
p2=p1(1-u) + (1-p1) v
p3=p2(1-u) + (1-p2) v
pi=pi-1(1-u)+(1-pi-1) v
at equilibrium this will be p = v / (u+v) and q = u/(u+v)
Because the forward mutation rates are rather small: around 10-8 mutations per generation and site for DNA, for microsatellite repeats more in the range of 10-5 by insepcting the above recurrence equations we can see that the change of allele frequencies will be slow from one generation to the next. We may need to wait millions of generations until the equilibirum of forward and backward mutation is reached.
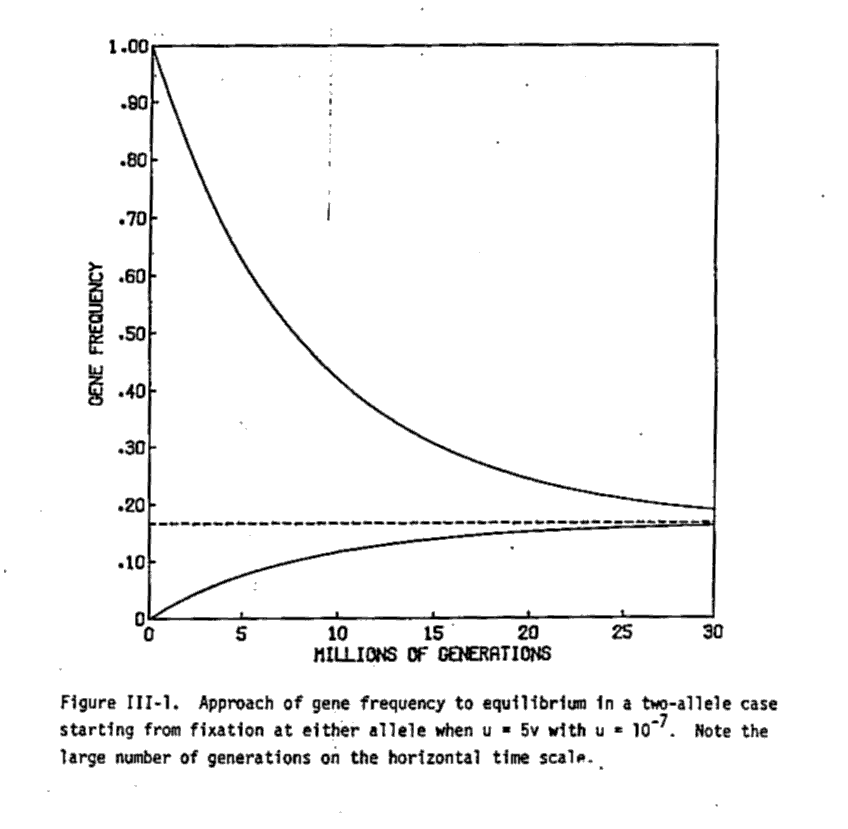
Some object that natural selection cannot work because on a extremely tiny fraction of sequences at a locus will be eve nmarginally functional. Mutants will almost always be totally nonfunctional. This objection is invalid because mutants do not reach random sequences but neighbors of functional ones. These neighbors are much more likely to be functional.
An analogy would be a workd game were we change one letter at a time, e.g. from WORD to GENE
in 4 steps and every intermediate still is an English word. There are 264=456,976 possible four-letter words of course most of them are gibberish like BGXU. Like meaningful words functional genes are clustered in clumps that are close to each other in sequence space.
The sequence space is very big, with 1000 bp, there are 41000 which is roughly 10602 sequences. Each sequence has 3000 neighbors one mutation away. Two random sequences with a length of 1000 bp are only about 750 steps apart.
When mutations are deleterious, natural selection will oppose mutation and will hold the deleterious mutants at low numbers.
Assume that we have a recessive deleterious mutant whose fitnesses are
| AA | Aa | aa | |
| Fitness | 1 | 1 | 1-s |
When the frequency of that deleterious allele a is q, in a large population of N individuals we expct Nq2 homozygotes, a fraction s of whom die [this makes it easier for our calculations]. Each death eliminates two copies of a. So the total loss of a copies is 2Nq2s per generation. 2Nu new copies of a arise by mutation if the mutation rate is u. Equating these we get the equilibrium frequency
This can be very low. For example, if we use a standard mutation rate u=10-8 and s=0.01 we get a equilibrium frequency of q=0.001.
| AA | Aa | aa | |
| Fitness | 1 | 1-hs | 1-s |