Migration is the process when individuals or their gametes move from one population to another. Other related terms: Gene flow, dispersal.
Migration is equalizing the allele frequencies in the different populations. This averaging is a weighted average. A population that consists of 80% individuals from population I and 20% from population II will have a allele frequency which is 0.8 pI + 0.2 pII, where pI and pII are the allele frequencies in populations I and II. If they have allele frequencies 0.9 and 0.3 for allele A the admixed population will have a p = 0.8(0.9) + 0.2(0.3) = 0.78.
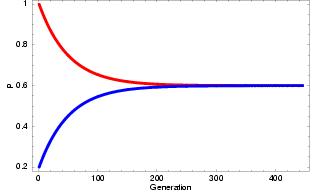
Continued migration between populations will bring their frequencies closer and closer to each other, at a rate that is controlled by the migration rates. In the end their gene frequencies will be expected to be the same as if all the gene copies at that locus would come from the same big gene pool. In the example we have two population with frequences p1 = p1(A)=1.0 and p2(A)=0.2, the migration rate is m=0.01. In each generation the frequency of p1(A) will change by the addition of alleles from population 2 and the loss of alleles that move to population 2:
p(1)1 = p(0)1(1-m) + p(0)2m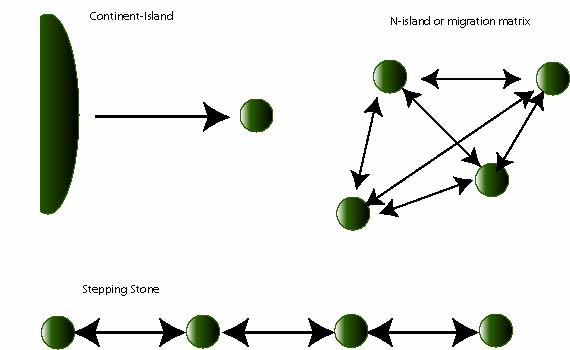
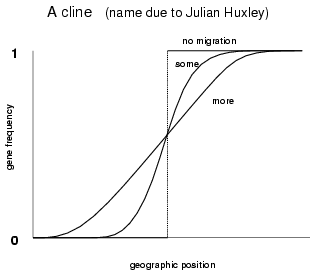
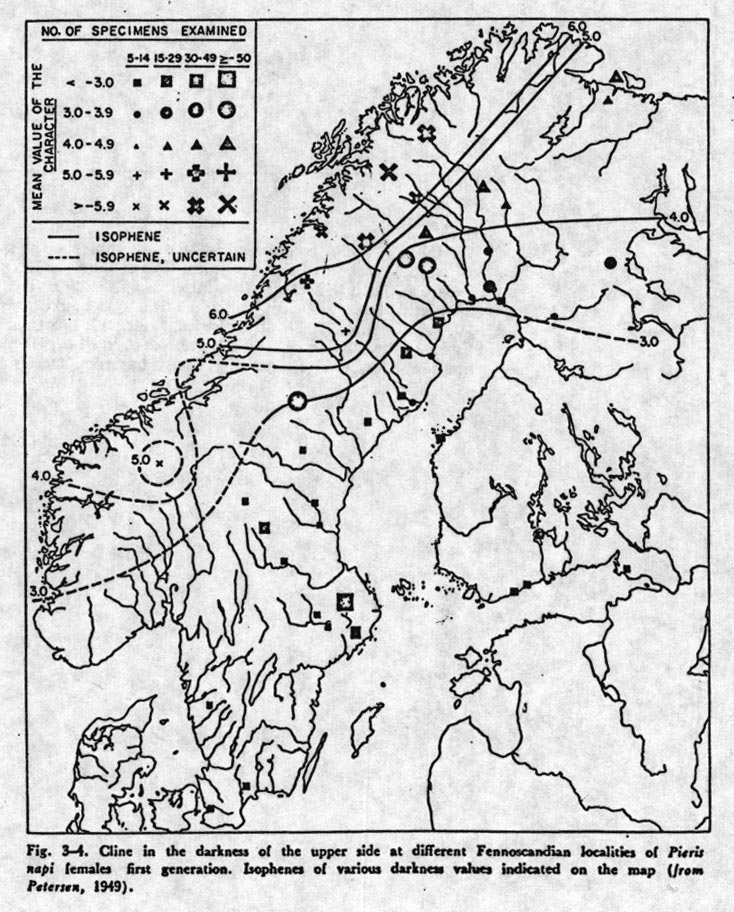
Natural selection and genetic drift can cause allele frequencies in different populations to differ. When there is a geographic continuum such as North-South, if natural selections differs in these geographic regions a pattern of smooth change of gene frequency (a cline) can be set up. It is not easy to tell from a cline's shape whether the selection regime changes smoothly as one goes from South to North, or whether it changes abruptly at an environmental boundary. Migration will create a smooth pattern of gene frequency in either case.
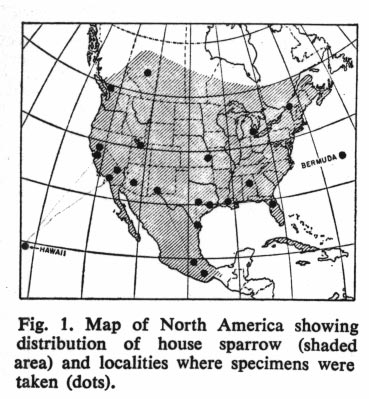
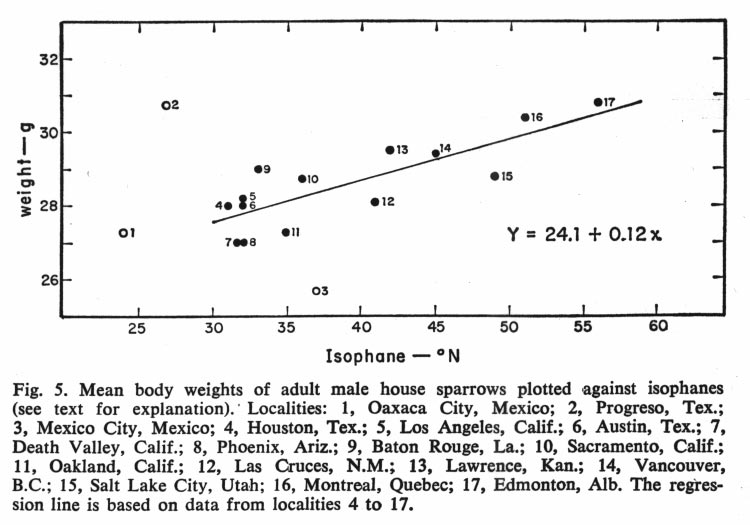
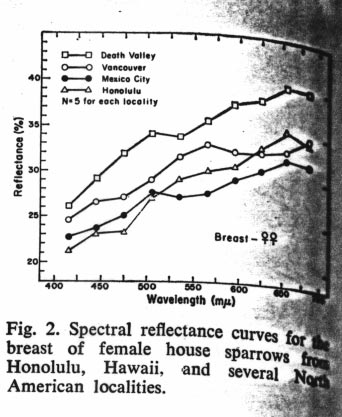
Johnston and Selander (Science 144: 548-550 1964) studied geographical variation in house sparrows (Passer domesticus) in North America. They have differentiated quite a bit geographically since the introduction in the mid-1800's. They follow the well-known "laws" of geographic differentiation in northern regions: they are larger in northern areas (Bergman's rule), have relatively shorter appendages in northern areas (Allen's rule), and are darker in northern areas (Glogler's rule).

|
|
 |
 |
 |
|
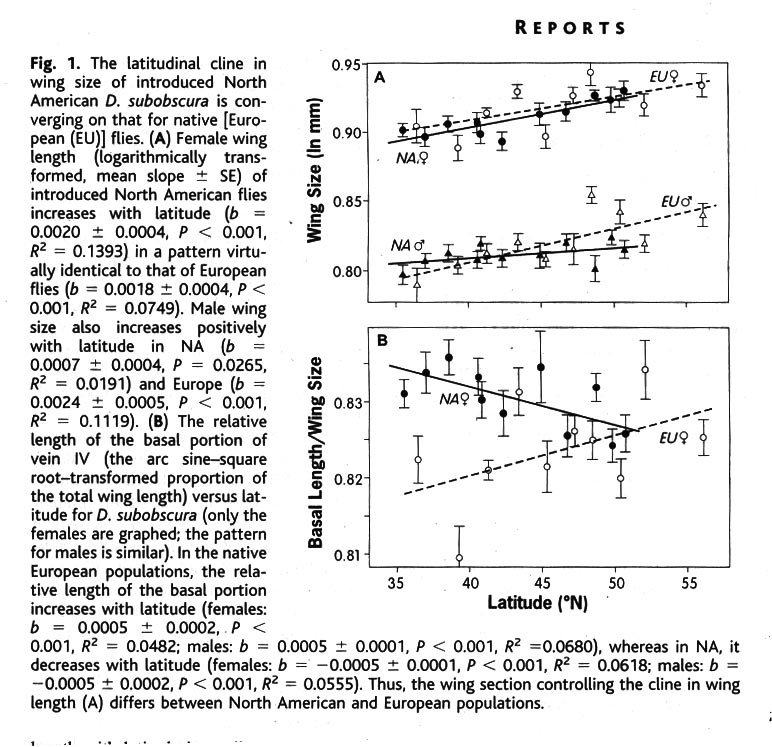
Wing length in Drosophila subobscura follows also an Equatorial-Pole cline (Huey et al. Science 287: 308-309 2000). The recently into the Americas introduced flies develop the same cline structure as in Europe. On both continents the have larger wings in the North, but this is achieved through different paths, on one continent it is an enlargement of the basal part of wing cell IV, and on the other it is the distal part is enlarged.
 |
|
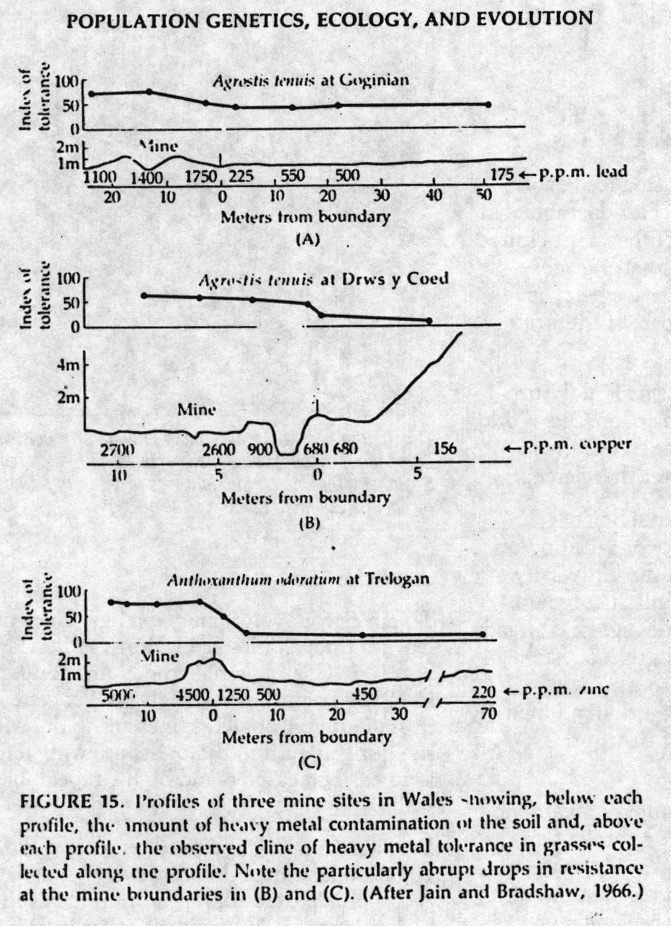
Antonovics, Bradshaw, and Turner (Advances in Ecological Research 7: 1-85 1971) examined heavy metal tolerance in plants growing in Welsh mines. Plants from within the mine area showed higher tolerance to the metal produced by that mine (and spread around in its soil due the dumping of tailings). The tolerance declines smoothly as one reaches the boundary of the mine. This was assessed by testing the descendants of the plants in common gardens, so these are almost certainly genetic clines.