Mitosis & Centrosomes
SUMMARY:
Dividing a eukaryotic cell in two is not trivial. This page gives information beyond what is normally discussed and is meant to be a supplement to a normal course of instruction.
The centrosome is a neglected cell organelle, yet is absolutely essential for eukaryotic cell division and cell structural organization.
Calcium - usually in the Ca+2 form. Calcium plays a crucial factor on cell signaling. Certain proteins react to the presence of calcium to turn on and off other proteins in the cell. Calcium concentrations are critical to the entire mitotic cycle. Calcium is released into the cytoplasm at the correct time from internal stores in vesicles associated with the mitotic apparatus.
Centrosome - provides the organizing matrix for keeping all the players in formation. See
Centrosome below.
Chromosomes - condensed DNA, carrying the genetic information of the cell. It is essential that the right chromosomes go to each half of the dividing cell.
Cytoplasm - a lot of debate as to whether the cytoplasm is liquid or gel, or some combination. In this case we refer to the ions, proteins and other cell components in solution around the mitotic apparatus.
Kineticore - the attachment site on the chromosome whereby the microtubules connect with the chromosomes, allowing them to be pulled to the poles.
Microtubules - multi unit structure built from a globular protein called tubulin.
Nuclear Envelope - the porous membrane surrounding the nucleus most of the cell cycle. The nuclear envelope provides a way of separating what happens in the nucleus from what happens in the cytoplasm.
Proteins - built from the 23 amino acids into polymers of astounding diversity. Proteins offer catalytic activity as enzymes, as structure components, as ion channels, and a host of other uses.
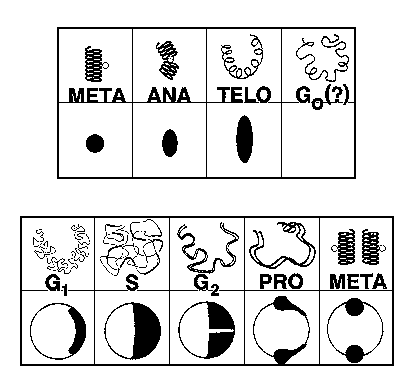
G0 – the stasis state of the unfertilized egg, waiting for the signal to activate. G0 does not exist in normal somatic cells.
G1 – normally referred to as the "growth" phase in somatic cells. The DNA is "quiet", but the cell is adding bulk, reproducing mitochondria and other structures.
S – the time of DNA synthesis. (also centrosome synthesis)
G2 – DNA synthesis is complete, waiting for the signal to start mitosis.
Prophase – mitosis is initiated, chromosomes are condensing and attaching to the centrosome via the microtubules and kineticores.
Metaphase – the chromosomes are lined up and sorted out, in a "plate" between the two centrosomes
Anaphase – the chromosomes are moved to the two poles of the mitotic apparatus set up by the two centrosomes.
Telophase – the physical dividing of the cell itself. Nuclear envelopes form around the two new nuclei.
The following illustrations show the timing relationships of the chromosomes (top) and the centrosomes (bottom).
Mitosis is not a static event. The best way to see and understand the relationships between all of the components is to add the TIME dimension. The following are links to animations done with the help of Dr. Daniel Mazia:
- Chromosome and Centrosome Cycle shows just the relationship between the chromosomes and the centrosome.
- Fertilization - shows the first two divisions in a developing sea urchin embryo.
- Unequal - shows how an unequal cell division can happen by an unequal division of the centrosome.
- Insect - in the fruit fly egg, mitosis is different. The egg is too big to divide at each division and fruit flies are protostomes. They use spiral cleavage instead of the radial cleavage as seen in sea urchins and humans.
- Cells - A cell must grow in order to divide, but this does not occur at the same time in the egg/embryo and in the somatic cell (body cell).
The centrosome is more than an organizing center for mitosis, it is quite possibly the structural organizing center for the entire cell. In spite of its importance, no one really knows what it is made of. There are proteins associated with the centrosome and these have been isolated and characterized, but the actual structural component has not been identified. Like the mitochondria, the chloroplast, and other cell components, the centrosome may be a remnant of an earlier symbiotic relationship with a prokaryotic organism.
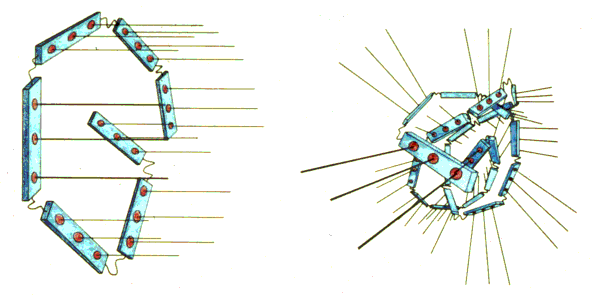
Recent work has lead to theories as to how the centrosome organizes microtubules.
The red buttons are the initiation points for microtubule elongation. There appears to be two structural components to the centrosome, the light blue core and the "linker" component between the core pieces. By arranging the centrosome components into different patterns, different arrays of microtubules can be achieved. Shown here are the "brush" pattern and the "ball" pattern, respectively.
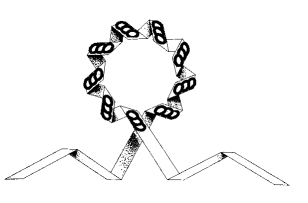
It may even be that the centrosome is the organizing center for cilia and flagella. By twisting the centrosome in the right way it is possible to get the standard array of microtubule initiators.
The string like centrosome can assume almost any shape, from the ball like metaphase "pole" shape to the more elongate streak and telophase shapes seen. The last shape shown has only been seen in the laboratory and not in living cells.