Morphology
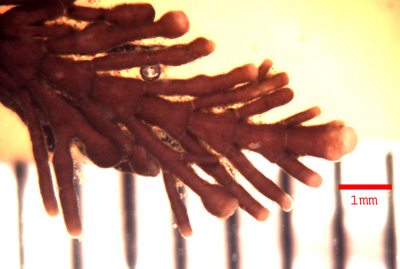
View of a Corallina vancouveriensis terminal thallus. Note branching is off the distal ends of the intergenicula. White lines between intergenicula are genicula. White bulbous portions at the ends are areas of new growth.
Backlighting of genicula (white and off white spaces) and intergenicula (darkened areas).
Longitudinal section through two intergenicula and two genicula. Genicula are denoted by the red bars beneath them. Intervening segments are the calcified intergenicula.
Longitudinal section of a geniculum. Note its clarity, a result of its having been decalcified.
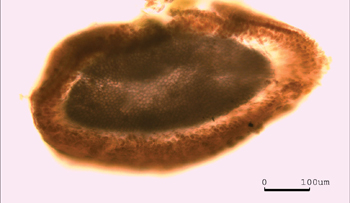
Cross section through an intergenicula of C. vancouveriensis.
|
This red alga has densely branched dark violet to purplish fronds that can be up to 14 cm in length. However, at the more Southern parts of its range it tends to be shorter and have a reduced length-width ratio (Johansen 1976, Riosmena- Rodríguez & Siqueiros-Beltrones 1995). It is epilithic and typically forms clumps off rock faces or is found erect in tide pools. The thallus is composed of genicula* and intergenicula with the latter having square dimensions with each side ~1mm in length (Johansen 1976). Most intergenicula bear 2-5 adaxially appressed branches and those on the main axis are in the shape of an inverted arrow with a narrow basal portion and expanded distal portion (Johansen 1976, Riosmena- Rodríguez & Siqueiros-Beltrones 1995). The final two intergenicula (ultimate and penultimate) are on occasion broad and conceptacles can grow as large as 500 μm in outer diameter (Johansen 1976). Morphology of this alga can also change on a local scale. In high energy zones the thalli are denser and their axes become flatter the more distal they are compared to forms found in lower energy settings (Riosmena- Rodríguez & Siqueiros-Beltrones 1995). C. vancouveriensis can grow as either individual thalli or dense turfs (Taylor & Hay 1984). This differentiation is correlated with its association with the sea anemone Anthopleura elegantissima (Taylor 1985). See Ecology for more details. In shallower water there is an increased amount of tier branching (Riosmena- Rodríguez & Siqueiros-Beltrones 1995). Like other coralline red algae, this species has pit plugs lacking cap membranes and has 2-layered plug caps that have their outer layers domed (van den Hoek et al 1995). * - Genicula and intergenicula are a hallmark of the coralline algae. Their purpose appears to be in providing both rigidity (intergenicula) and flexibility (genicula) in high wave action environments. For more information on the biomechanics of these structures see Martone 2006. Johansen, H. W. 1976. Family Corallinaceae. In. A. Abbott and G.J. Hollenberg (eds), Marine algae of California. Stanford Univ. Press, California, 379-419pp. Martone, P. T. 2006. Size, strength and allometry of joints in the articulated coralline Calliarthron. Journal of Experimental Biology. 209, 1678-1689. Riosmena-Rodríguez, R. & Siqueiros-Beltrones, D. A. 1995. Morphology and distribution of Corallina vancouverensis (Corallinales, Rhodophyta) in northwest Mexico. Ciencias Marinas 21, 187-199. Taylor, P.R. & Hay, M. E. 1984. Functional morphology of intertidal seaweeds: adaptive significance of aggregate vs. solitary forms. Marine Ecology Progress Series. 18, 295-302. Taylor, P. R. 1985. The influence of sea anemones on the morphology and productivity of two intertidal seaweeds. Journal of Phycology. 21, 335-340.. |