RESEARCH
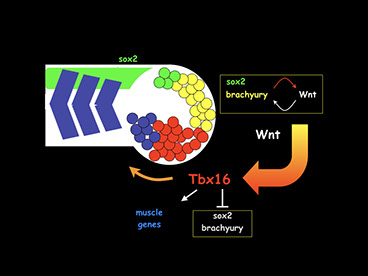
Vertebrates undergo an amazing and rapid process of growth during early development to form the trunk and tail of the body. Zebrafish, like all vertebrates, grow from the neck down by progressively adding cells from a progenitor population, which contributes cells to both the mesodermal (muscle and bone) and neural (spinal cord) tissues, located at the most posterior end of the embryo. A major interest in the lab is to understand the mechanisms that regulate this process. To date, a principal focus has been on the signaling factors that pattern and regulate the mesodermal progenitors, including Wnts, Fgfs, Bmps, Nodals and retinoic acid, and their interaction with the transcription factors that regulate the progenitors, particularly members of the T-box family. We showed (Martin and Kimelman, 2012) that the posterior of the body forms from a bipotential progenitor population, which produces both the neurons and mesoderm, with Wnt signaling determining which fate the progenitors will adopt. Wnt expression is controlled by the T-box transcription factor Brachyury (called No tail in zebrafish) in an autoregulatory loop (Martin and Kimelman, 2008), which sustains the progenitors in a multipotent state. As cells exit the progenitor state, many of them turn on another T-box transcription factor, Tbx16/Spadetail, which locks cells in the mesodermal state by turning off progenitor genes such as brachyury and sox2, and turning on the early mesodermal genes (Bouldin et al., 2015).
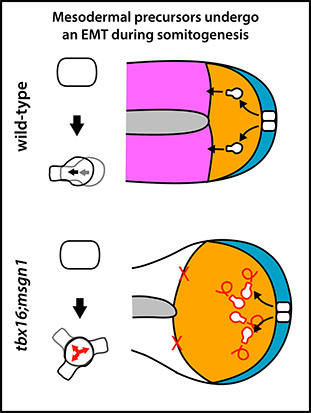
A major current focus is on the regulation of morphogenesis of the mesodermal cells. As a multipotent progenitor cell begins to differentiate as mesoderm, it undergoes an epithelial to mesenchymal (EMT)-like transition. We found that Tbx16 plays a key role in this process (Row et al., 2011). In tbx16 (spadetail) mutants, cells begin the transition but get stuck in an intermediate highly blebbing state during the gastrula stages, preventing them from completing the morphogenetic transformation, resulting in partially differentiated cells trapped at the most posterior end of the body and a failure to form normal muscle. These results reveal a novel intermediate step in the EMT process. Using a novel explant system we recently developed, we now have been able to image this process during the somite forming stages (Manning and Kimelman, 2015). While Tbx16 continues to play a major role in morphogenesis, surprisingly its function changes during these later stages and now it is needed for directed migration after the EMT. These results show the essential, but changing role of Tbx16 in regulating embryonic morphogenesis.