LIFE HISTORY
|
Schematic illustrating the two methods of reproduction in seagrasses.
|
|
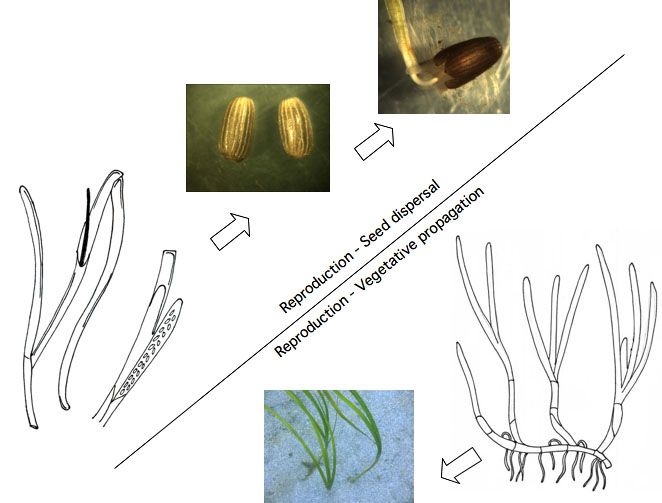
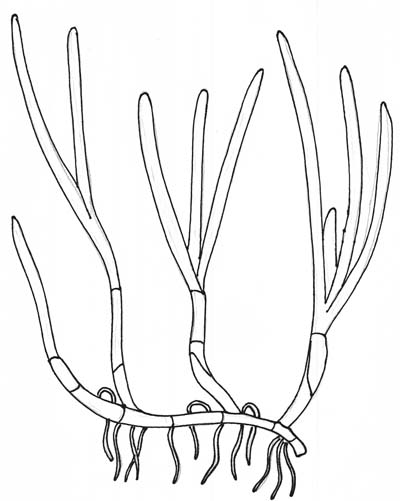
Vegetative Growth
Seed Dispersal
In contrast to terrestrial monocotyledons, the morphology and biology of seagrasses has been modified to complete their life cycle in the marine environment.
Vegetative propagation
Seagrasses commonly reproduce by means of vegetative propagation, where the plant sends out rhizomes (horizontal, underground stems) that send up new shoots of seagrass. In this way, eelgrass spreads quickly over a small area.
Generative growth - seed dispersal
The second method is to spread by seed dispersal, which is a slower method of reproduction, but can broadcast offspring over a greater distance (Orth et al., 2006).
Most seagrasses are perennial, however there are a few exceptions. iZ.marina is both an annual and a perennial (Hemminga & Duarte, 2000). Z.marina depends completely on generative growth (i.e. recruitment of seeds) rather than generative and vegetative growth (Orth et al., 2006).
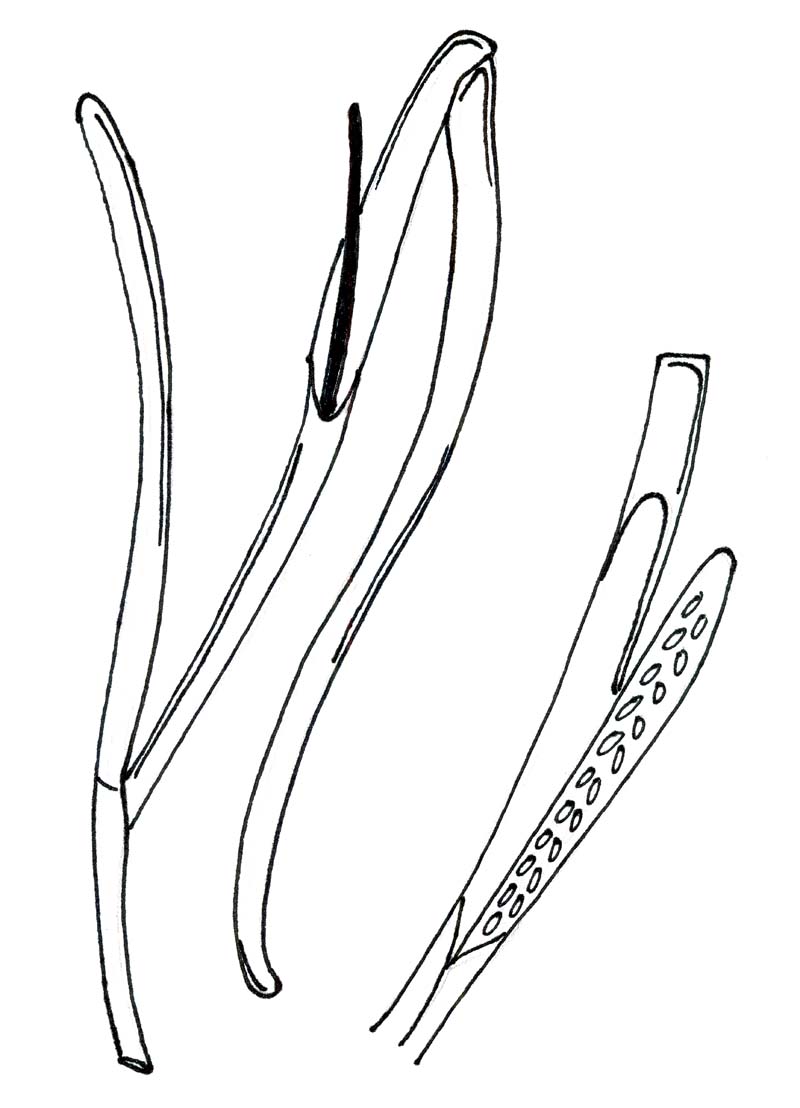
Generative shoots (inflorescent shoots) initiate as terminal or lateral branches at rhizome nodes. A mature reproductive shoot-bearing node usually appears a few rhizome nodes behind a regulative shoot bearing node of the extending rhizome (Hemminga & Duarte, 2000). It is important to note that most seagrass shoots rarely bear flowers or fruit, and the vast majority of shoots are lost before ever becoming reproductive.
Flowering of seagrasses is controlled by temperature and often occurs simultaneously across large spatial scales, when irradiance improves and water quality increases (Hemminga & Duarte, 2000).Flowers are formed on flattened, spike-like axis known as the spadix, and they are enveloped by a modified open leaf sheath – spatial sheath (Orth et al., 2006). Mature stamens of seagrass form longitudinal slits and release their pollen into the water, where it travels with the current before it attaches to the stigma surface. Either at the water surface or below. Z.marina exhibits hydrophilous pollination, and both the pollen and the stigma are coated with an adhesive substance, which binds strongly upon contact (Hemminga & Duarte, 2000).

Photograph of dry Z. marina seeds.

Photograph showing the germination of a Z. marina seedling
Seeds
Seagrass seeds are achene with a sclerous pericarp and a hard, fibrous endocarp as a seed ‘coat’ (Orth et al., 2006). Seed morphology is species specific, with variation in shape (hooks vs. beads) and size (implications for dispersal and fitness). Overall morphology is dependent on the environmental stressors present.
Dynamics & Dispersal
Seeds are typically negatively buoyant and do not disperse very far. Once in the sediment, Z. marina seeds can remain dormant for 1-2 months (Hemminga & Duarte, 2000).
Studies on Z. marina populations have shown that 95% of seeds are retained within 30 m from the source (Ruckelshaus, 1996). There is also evidence to suggest that seagrass seeds experience high mortality due to many factors, including lack of viability, physical damage, export to unsuitable areas, burial and predation (Ruckelshaus, 1996).
© Jessica Smith 2008
Hemminga. M, C. Duarte. (2000). Seagrass Ecology. Cambridge. United Kingdom.
Larkum.A, R.Orth, C.Duarte. (2006). Seagrasses: Biology, Ecology and Conservation. Springer.
Orth, R, M. Harwell, G. Inglis (2006) In: Larkum.A, R.Orth, C.Duarte. (2006). Seagrasses: Biology, Ecology and Conservation. Springer.
Ruckelshaus, M. (1996). Estimation of genetic neighbourhood parameters from pollen and seed dispersal in the marine angiosperm Zostera marina, L. Evolution. 50: 856-64.